
Abstract
Inhibitory chemical compounds (ICCs) as by-products and acidic conditions during plant biomass pretreatment were inhibited microbial fermentation. This study aimed to select and characterize the inhibitory and acid-resistant chemical compounds of unique yeasts capable of producing xylitol. Meyerozyma guilliermondii InaCCY65 was selected as a potential candidate for bioconversion of lignocellulose to xylitol. This strain can grow in various hydrolysates in acidic pH and still produce xylitol. The optimum temperature and pH of InaCCY65 for producing xylitol are 30°C and pH 4.5. This strain was tolerating ICC and capable of detoxifying ICCs. The results will provide new technology to produce xylitol with yeast that are robust and tolerant to ICC and acidic conditions.
Keywords: acid tolerance, fermentation, inhibitor compounds, lignocellulose, tolerance, xylitol; xylose, yeast
Outline
1. Introduction
Xylitol (C5H12O5) is a sugar-containing alcohol functional groups favored as a sucrose alternative due to its similar sweetness level (0.8 to 1.0) and lower caloric content of 2.4 calories per gram, as opposed to sucrose's 4 calories per gram1). Its consumption is linked to various health benefits, notably oral health, as it hampers microbial growth, thus reducing or preventing tooth decay2). Additionally, xylitol shows promise in managing conditions such as diabetes, hemolytic anemia, inflammatory disorders, and colon diseases while also serving as a preventive measure against respiratory infections, acute otitis media, and osteoporosis3). Over recent decades, the extensive utilization of xylitol across food and pharmaceutical sectors has surged, consequently broadening its market reach4). At an industrial scale, xylitol production involves chemically reducing D-xylose, typically employing a nickel catalyst at high temperatures, yielding a conversion efficiency of approximately 50-60%5). This chemical process entails temperatures ranging from 80-140°C and pressures exceeding 50 atm and necessitates subsequent separation and purification steps6). The involvement of a catalyst escalates production costs, contributing to the overall expense of the chemical process7).
Biological processes are considered efficient and can potentially reduce the cost of xylitol production8). Bioconversion of xylose to xylitol involves hydrolysate containing xylose and yeast as the conversion agent. In pursuit of sustainable practices, the utilization of biomass for chemical compound production has evolved into the concept of a biorefinery plant9). Through an integrated technological approach, plant biomass can be effectively converted into various valuable compounds, thereby minimizing waste generation10). Some plant biomass has the potential as a substrate for biofuel and bioproducts, such as corn cob11), sugarcane12), rice straw13), and others. Plant biomass typically comprises cellulose, hemicellulose, and lignin14), with hemicellulose containing five neutral sugars: glucose, mannose, and galactose (hexosan), as well as xylose and arabinose (pentosan) being the predominant components15). The hydrolysis of hemicellulose presents the opportunity to obtain xylose, which, derived from plant biomass, acts as a potential raw source for xylitol16). However, the depolymerization process usually results in an accumulation of growth-inhibitory compounds, such as 5-hydroxymethylfurfural (HMF), furfural, acetic acid, and phenolic compounds17).
The microbial bioconversion of xylose into xylitol boasts an impressive conversion efficiency ranging from 65% to 85%18). Yeast, among various microorganisms, stands out as particularly proficient in xylitol production19). Opting for yeast-mediated bioconversion presents an attractive alternative to industrial chemical processes. This approach offers notable advantages, and it reduces energy consumption and a more sustainable, and eco-friendly profile20). Several yeasts have been reported as the cell factory for xylitol, such as Candida tropicalis21), C. sojae22), Wickerhamomyces anomalus23), C. guilliermondii24), Debaryomyces hansenii25), Kluyveromyces marxianus26), Meyerozyma guilliermondii 27), and Clavispora lusitaniae28).
The biological synthesis of xylitol involves yeast-mediated bioconversion of xylose, facilitated by the enzyme xylose reductase29). This process from plant biomass entails various stages, including pretreatment of the biomasses30), conversion of xylose monomers to xylitol using yeast8), followed by purification and crystallization to obtain the final xylitol product31).
Fermenting xylitol from lignocellulosic biomass conversion using yeast poses various challenges, particularly during the biomass pretreatment stage, where inhibitory compounds and low pH are produced, hindering yeast fermentation32). These inhibitory compounds disrupt yeast metabolism33). Despite yeast strains from the Candida genus being identified as superior xylitol producers34), Candida species are also known to cause various diseases (such as candida, endocarditis, vaginitis, meningitis, and endophthalmitis)35). In addition, pre-treatment of lignocellulosic biomass using acid produces sugars, ICCs and acidic conditions are also formed. These ICCs and acidic pH compounds affected to yeast metabolism36). Therefore, it is necessary to explore non-Candida yeast strains that are robust for xylitol production, and tolerant to ICCs and acidic pH. Thus, yeast is safe for food applications, and effective in the bioconversion of lignocellulose into xylitol. Hence, there's a need to explore non-Candida yeast strains for xylitol production, characterizing their yield, tolerance to inhibitors, and safety for food applications. This study intended to identify and assess the biotransformation of xylose from lignocellulosic biomass into xylitol using non-Candida yeast strains with high conversion yields and resistance to complex chemical inhibitors and acid pH.
Accessing the potential of non-conventional yeast in hemicellulose hydrolysate for xylitol is an important step for the development of sustainable biorefinery and economic models37). Therefore, in this study, ninety six yeast strains from the Indonesian Culture Collection (InaCC) were selected and screened on xylose, hydrolysates of corn cobs (CC), sugarcane leaves (SCT), and oil palm empty fruit bunches (OPEFB) in various pH. The selected strains were further characterized, focusing on the growth pattern of xylose sugar consumption, and xylitol production under Inhibitory Complex Compounds (ICCs) stress conditions. It is hoped that one potential strain that is resistant to low pH and ICCs compounds can be obtained, so that an efficient xylitol biorefinery from lignocellulosic waste can be developed.
2. Material and Methods
2.1. Yeast strain
This study involved the screening and evaluation of ninety-nine yeast strains obtained from the Indonesia Culture Collection (InaCC) at the National Research and Innovation Agency (BRIN). The subculturing and maintenance process of these strains were cultivated on Yeast Peptone Glucose (YPG) agar medium containing 10 g/L yeast extract, 20 g/L bacto peptone, 20 g/L glucose, and 15 g/L agar. The pre-culture was prepared with inoculated a single colony from the YPG plate into 12 mL of YPG liquid medium in a 100 mL Erlenmeyer flask. The culture was then incubated overnight at 30°C with agitation at 150 rpm.
2.2. Growth test of 96 strains of InaCC yeasts in xylose
In the initial screening phase, ninety-nine yeast strains sourced from InaCC were cultivated in Yeast Peptone Glucose (YPX) media. A seed culture of 100 µL was utilized to inoculate 1000 µL of fermentation medium in deep well plates, resulting in a 10% inoculation size. The yeasts were then cultured in 1000 µL of YPX media comprising 10 g/L yeast extract, 20 g/L peptone, and 20 g/L xylose. Incubation process at 30°C, with agitation at 1400 rpm for 24 hours using an incubator-shaker (Bio Shaker MBR-022UP, Titec, Japan), all experiments were performed in triplicate. The cell growth was analyzed using a spectrophotometer at a wavelength of 600 nm, while product fermentation analysis was carried out using HPLC. The non-Candida yeast strain with the highest xylitol production was selected for further investigation.
2.3. Growth test of selected non-Candida and xylitol-producing InaCC yeasts in various biomass hydrolysates and at several pH
Hydrolysates of corn cob (CC), sugarcane trash (SCT), and oil palm empty fruit bunch (OPEFB) underwent hydrolysis using 1.8% (w/v) maleic acid and were then subjected to microwave heating for 60 minutes30). Subsequently, the hydrolysate was adjusted to pH levels of 2.0, 3.0, 4.0, 5.0, and 6.0. After sterilization using a 0.45 µm filtration membrane, the hydrolysate was utilized for xylitol fermentation. Eleven yeast strains selected for the study were cultivated in YM 10X media containing predetermined mineral concentrations at various pH levels. The YM medium composition, consisting of 10 g/L yeast extract, 10 g (NH4)2SO4, 2.4 g KH2PO4, 0.2 g MgSO4, and 0.3 g CaCl2, served as the fermentation medium for xylitol23). The YM medium was sterilized at 115°C for 15 minutes. The SCT, CC, and OPEFB hydrolysates were introduced into the fermentation medium as carbon and nitrogen sources. To initiate fermentation in the YM medium, the initial cell concentration aimed to achieve an optical density of 0.3 at a wavelength of 600 nm. Fermentation was carried out in 2000 µL deep well plates, each containing 100 µL of seed culture, YM 10X medium, and hydrolysate, with a total culture volume of 1000 µL. The cultures were then incubated at 30°C with agitation at 1400 rpm for 24 hours using an incubator-shaker (Bio Shaker MBR-022UP, Titec, Japan). All experiments were performed in triplicate. The cell growth was analyzed by using a spectrophotometer at a wavelength of 600 nm, and product fermentation analysis was performed using HPLC. The non-Candida yeast strain with the highest xylitol production was selected for further investigation.
2.4. Growth test of InaCCY65 strain in pH and temperatures
The optimal pH and temperatures for the InaCCY65 strain were determined through growth tests and xylitol production assessments conducted across varying temperature and pH levels. Initially, the growth of the InaCCY65 strain was evaluated at pH 4.5 under different temperature conditions, ranging from 25°C to 40°C. The cultures were incubated for 30 hours at 150 rpm. Following the identification of the temperature yielding the highest xylitol concentration, subsequent growth trials were conducted at various pH levels. The YPX medium was adjusted to pH values of 2.0, 3.0, 4.0, 5.0, and 6.0, with incubation at 30°C. The fermentation process of InaCCY65 yeast strain seed was inoculated into a 100 mL Erlenmeyer flask containing 12 mL of YPX medium. The initial cell concentration aimed to achieve an optical density of 0.3 at a wavelength of 600 nm. All experiments were carried out in triplicate, and cell growth was monitored using a spectrophotometer at a wavelength of 600 nm. Product analysis was conducted using HPLC. The yeast strain demonstrating the highest xylitol production was selected for further investigation.
2.5. Fermentation of InaCCY65 strain in various xylose concentrations
The optimal concentration of xylose during early fermentation was investigated using a YPX medium with varying xylose concentrations. The InaCCY65 strain was cultured in 12 mL of YPX media containing 5, 20, 40, 50, 60, and 70 g/L xylose. Fermentation was carried out at 150 rpm and 30°C for 30 hours. The initial optical density of yeast at OD600 was set to 0.3 at the start of fermentation. All experiments were performed in triplicate, and the cell growth was monitored using a spectrophotometer at a wavelength of 600 nm. Fermentation products, including xylose consumption and xylitol production, were analyzed using HPLC.
2.6. Fermentation of InaCCY65 strain in medium containing inhibitor complex compounds (ICCs)
The batch cultivations were conducted in 100 mL Erlenmeyer flasks at 30°C and 150 rpm. In this investigation, YPX medium served as the control medium without inhibitors. Another medium, YPX medium, added with inhibitory compounds, including 60 mM furfural, 10 mM 5-HMF, 5 mM levulinic acid, 60 mM acetic acid, and 30 mM formic acid, was designated as the inhibitory medium (YPXi). The YPXi medium was then diluted with YPX medium at 0.25-fold (0.75YPX+0.25YPXi), 0.5-fold (0.5YPX+0.5YPXi), and 0.75-fold (0.25YPX+0.75YPXi) concentrations. The concentrations of ICCs in the YPXi medium were tuned to previously reported values that effectively inhibit the development of most yeast cells38). The batch cultivations were carried out in 100 mL Erlenmeyer flasks at 30°C and 150 rpm in an orbital shaker incubator for 30 hours. A pre-culture was utilized to inoculate 12 mL of fermentation medium into 100 mL Erlenmeyer flasks, aiming for a cell culture concentration of OD600 = 0.3. Samples were collected every 3 hours throughout the fermentation process. The samples were centrifuged at 6000 g for 5 minutes, and 400 µL of supernatant was transferred into vials for HPLC analysis. The cell growth was monitored by measuring OD600 using a UV–VIS Spectrophotometer (Shimadzu, Tokyo).
2.7. Fermentation product analysis
Cell growth was monitored by measuring the optical density at 600 nm (OD600) using a UV–VIS Spectrophotometer, specifically the UV mini-1240 model manufactured by Shimadzu in Tokyo. Xylose and by-products, including acetic acid, formic acid, levulinic acid, and xylitol, were analyzed using an HPLC system (LC20AB, Shimadzu, Tokyo) equipped with a COREGEL-87H3 column (7.8 mm I.D. 300 mm, Transgenomic Inc., New Haven, CT) and regulated at 80°C. The samples were eluted at a flow rate of 0.6 mL/min using 5 mM H2SO4 as the mobile phase. Additionally, the HPLC column Amminex HPX-87H (BioRad Labs. Inc., Hercules, CA) was utilized for assessing the concentration of inhibitors, predominantly furfural and 5-HMF, under similar conditions.
3. Result and discussion
3.1. Screening of xylitol-producing yeast in xylose substrate
To identify yeast strains with superior xylitol production capabilities, a selection was made from 99 InaCC yeast isolates, including Candida, Meyerozyma, Pichia, Trichosporon, Hyphopichia, Wickerhamomyces, and Kodama. Each yeast strain is identified by a numerical code preceding its name. The colored circles denote the xylitol concentration produced by each yeast strain, with red asterisks highlighting the strains chosen for further investigation (Figure 1). Although most of the screened isolates belonged to the Candida genus, none were identified as C. tropicalis, as this species is known to potentially cause human diseases39).
Among the 99 yeast strains tested, Candida jaronii (InaCCY78) emerged as the top xylitol producer, yielding 24.7 g/L. Additionally, among non-Candida strains, 11 demonstrated xylitol production exceeding 20 g/L, while five strains produced between 15 and 20 g/L. Four strains yielded xylitol concentrations ranging from 20 to 25 g/L. The screening results were found seven strains exhibited xylitol production exceeding 25 g/L (refer to Figure 1). The objective analysis identifies non-Candida yeast strains capable of utilizing agricultural waste hydrolysate and possessing robust xylitol production capabilities (as illustrated in Figure 1); we selected 11 strains for further investigation. We utilized six yeast genera identified from the initial selection results: Cyberlindnera, Hyphopichia, Wickerhamomyces, Rhodosporidium, Meyerozyma, and Pichia. Several of these genera have been documented in various publications for their ability to produce xylitol. For instance, Cyberlindnera fabiani, isolated from lignocellulosic material, has been reported to exhibit xylitol-producing capabilities40). Similarly, studies by41) have indicated that Cyberlindnera strains can produce xylitol from sugarcane trash. Likewise, Hyphopicia has been documented as capable of xylitol production42). Wickerhamomyces anomalus has also been recognized as
a xylitol-producing yeast23), particularly from sugarcane trash41). Rhodosporidium turoloides has been reported for its ability to produce lipids and xylitol from water hyacinth
(Eichhornia crassipes) hydrolysate43). Meyerozyma has

shown xylitol production potential from various hydrolysates, including rice straw44) and sugarcane trash45). Furthermore, Pichia has been demonstrated to produce xylitol from corncob46), sugarcane trash, and olive pits hydrolysates47).
Previous research has highlighted the yeast strain we selected, which several researchers have previously documented. However, exploring its xylitol production capabilities across various hydrolysates and pH levels remains intriguing. Hence, we chose 11 yeast strains to culture in corncob, sugarcane trash, and OPEFB hydrolysates. The results, including cell growth analysis and the concentrations of xylose and xylitol resulting from fermentation, are depicted in Figure 2.
Yeast strains capable of yielding high xylitol concentrations have long been of interest. Carneiro et al. identified six strains- Meyerozyma spp., M. guilliermondii B12, M. guilliermondii A3, Spathaspora sp. JA1 and Wickerhamomyces anomalus 740-aree the most proficient in utilizing xylose. Depending on the substrate employed, these yeasts displayed a wide range of yield factors, from YP/S = 0.25 g/g to YP/S = 0.60 g/g, with M. guilliermondii B12 achieving the highest yield factor of YP/S = 0.60 g/g when utilizing bagasse hydrolysate42).
In our investigation, we observed that M. guilliermondii InaCC Y65, isolated from Ledok, Cepu, Central Java, Indonesia, produced approximately 5 g/L of xylitol using xylose as the medium. Despite other strains, InaCC Y65 underwent further examination to explore the potential utilization of various agricultural by-products for xylitol production.
3.2. Screening of xylitol-producing yeast in lignocellulosic hydrolysates and pH
In pursuit of superior strains capable of utilizing agricultural residues for xylitol production, a hydrolysate was derived from three abundant agricultural by-products: sugarcane trash, corncobs, and oil palm empty fruit bunch. The material underwent immersion in sulfuric acid to achieve a pH range of 2 to 6. The resulting hydrolysate was then assessed for its xylose content. Subsequently, the substrate was inoculated with 11 selected isolates, specifically strains capable of producing xylitol in excess of 5 g/L. The outcomes of this selection process are depicted in Figure 2.
Figure 2 illustrates the varying xylitol production capacities of 11 yeast strains across corncob, sugarcane trash, and OPEFB hydrolysates at pH levels ranging from 2 to 6. The M. guilliermondii InaCC Y65 exhibited robust growth and xylose assimilation, yielding relatively high xylitol concentrations (4-5 g/L) from corncobs and sugarcane trash hydrolysates at pH 3-5. However, this strain produced approximately 3.0-3.9 g/L of xylitol when utilizing OPEFB hydrolysate in the medium at pH 5-6. Additionally, another strain from the same genus, M. caribbica InaCCY67, demonstrated xylitol production potential by hydrolyzing corncobs at pH 4 and sugarcane trash at pH 5. Consequently, M. guilliermondii InaCCY65 was selected for further xylitol production using lignocellulosic hydrolysate.
Furthermore, other tested strains, including Cyberlindnera fabiani InaCC Y34, Hyphopicia burtonii InaCC Y35 and InaCC Y44, Wickerhamomyces anomalus InaCC Y52, Rhodosporidium turoloides InaCC Y57, Hyphopicia burtonii Y66, Meyerozyma caribbia InaCC 67, 87, 88, also exhibited the capability to utilize corncob and sugarcane trash hydrolysates. This indicates that yeast strains with xylitol-producing potential are taxonomically diverse. To our knowledge, this study represents the first report of M. caribbica InaCC Y67, InaCC Y8, InaCC Y88, and Pichia silvicola InaCC Y95, which can assimilate corncobs and sugarcane trash hydrolysates for xylitol production.
We conducted a comparison of the YP/S values among different isolates across various pH hydrolysates, revealing that the InaCC Y65 isolate exhibited a higher value compared to other strains (see Table 1). InaCC Y65

demonstrated the ability to convert xylose from corncob and sugarcane trash hydrolysates into xylitol across a pH range of 2 to 6, respectively. Notably, the corncob hydrolysate at pH 4 and pH 5 exhibited the highest YP/S values, exceedingly approximately 0.5. The sugarcane trash hydrolysate at pH 3-5 displayed the highest YP/S value of approximately 0.4. Conversely, in the case of OPEFB, xylitol production by the InaCC Y65 isolate was only observed at pH 5.0, yielding a YP/S value of 0.1. This contrasts with Debaryomyces hansenii ITBCC R85, who demonstrated that xylitol formation only occurred at pH 5.0 after 24 hours of incubation, with a xylitol concentration of 0.047 g/L and a xylitol yield against xylose of 0.045 g/g48). These results suggest that InaCC Y65 exhibits a higher YP/S at OPEFB compared to D. hansenii ITBCC R85. Furthermore, the InaCC Y65 isolate demonstrated relative resistance and a broad pH range for xylitol production, spanning from pH 2 to 6.
Xylitol production is preferably reduced at higher pH levels, such as pH 6, due to its impact on the xylose transport system, thereby restricting xylitol formation within the fermentation solution and impeding its uptake by the cells. Conversely, at lower pH levels, such as pH 2 and 3, xylitol formation is also relatively limited. This phenomenon is attributed to disturbances in the redox balance of the bioconversion process, alterations in intracellular pH leading to a decline in the enzymatic reaction rate for xylitol synthesis, compromised yeast cell permeability, and the lack of assimilation of micronutrients owing to micronutrient precipitation47).
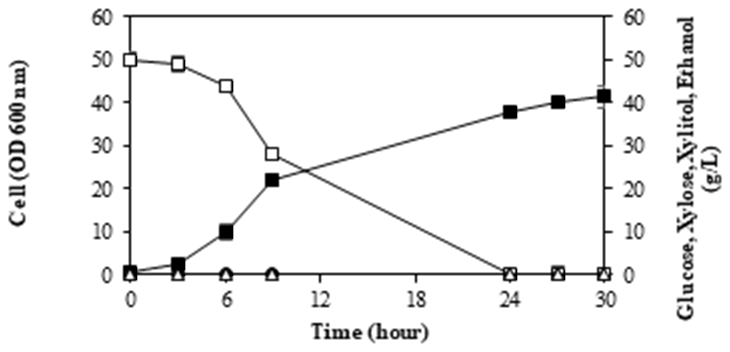
3.3. M. guilliermondii InaCCY65 in glucose-xylose as substrate
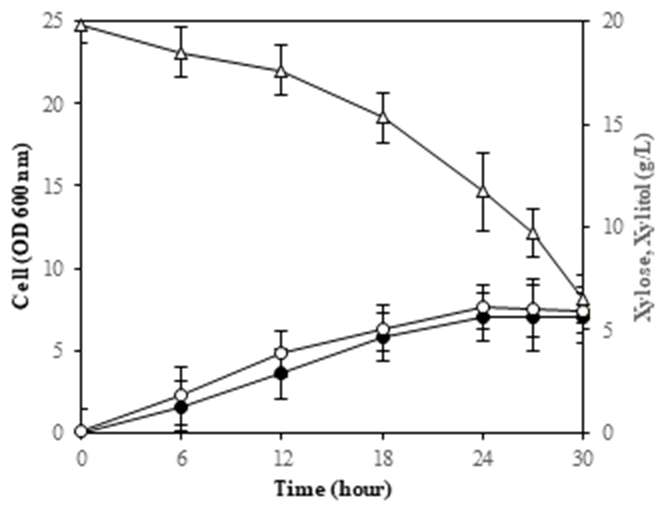
To investigate the effect of various carbon sources on the yeast's xylitol production capacity, glucose and xylose were used as culture mediums. The outcomes of the tests conducted using these two carbon sources are consolidated and illustrated in Figure 3.
The physiological traits of M. guilliermondii InaCCY65 were examined under both glucose and xylose conditions, where both sugars were utilized for biomass growth (Figure 3). Xylose consumption commenced only after glucose depletion in the medium containing both glucose and xylose. Xylitol production occurred subsequent to xylose assimilation and was exclusively observed in the presence of xylose in the medium; glucose was not utilized
Table 1: The YP/S of eleven yeast strain in a YP medium containing corncob, sugarcane trash, and OPEFB after incubation at 30oC during 24 h incubation (p < 0.05, Tukey’s test)
| pH | Strain Code of InaCC | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Y34 | Y35 | Y44 | Y52 | Y57 | Y65 | Y66 | Y67 | Y87 | Y88 | Y95 | |||
| Hydrolisate | Corncob | 2 | 0,1±0,12bcB | 0,2±0,15bcB | 0,1±0,11cB | 0,1±0,09cB | 0,2±0,11bcB | 0,2±0,15aB | 0,2±0,12bcB | 0,2±0,11abB | 0,1±0,10cB | 0,1±0,12cB | 0,1±0,11bcB |
| 3 | 0,2±0,11bcB | 0,2±0,11bcB | 0,1±0,15cB | 0,1±0,11cB | 0,1±0,12bcB | 0,3±0,14aB | 0,2±0,11bcB | 0,2±0,11abB | 0,1±0,12cB | 0,1±0,11cB | 0,1±0,14bcB | ||
| 4 | 0,2±0,12bcA | 0,2±0,11bcA | 0,1±0,14cA | 0,1±0,13cA | 0,2±0,12bcA | 0,5±0,12aA | 0,3±0,10bcA | 0,4±0,11abA | 0,1±0,12cA | 0,2±0,15cA | 0,2±0,11bcA | ||
| 5 | 0,2±0,14bcA | 0,2±0,12bcA | 0,2±0,11cA | 0,2±0,10cA | 0,2±0,09bcA | 0,5±0,09aA | 0,2±0,11bcA | 0,3±0,10abA | 0,2±0,11cA | 0,2±0,11cA | 0,2±0,10bcA | ||
| 6 | 0,1±10bcB | 0,1±0,11bcB | 0,1±0,12cB | 0,1±10cB | 0,1±0,12bcB | 0,2±0,11aB | 0,1±0,09bcB | 0,2±0,10abB | 0,1±0,10cB | 0,1±0,11cB | 0,2±0,11bcB | ||
| Sugarcane trash | 2 | 0,1±0,12abC | 0,1±0,11abC | 0,1±0,11bC | 0,1±0,13abC | 0,1±0,10abC | 0,2±0,09aC | 0,2±0,11abC | 0,2±0,13abC | 0,1±0,10abC | 0,2±0,11abC | 0,2±0,10abC | |
| 3 | 0,2±0,12abBC | 0,2±0,10abBC | 0,2±0,11bBC | 0,2±0,11abBC | 0,3±0,10abBC | 0,4±0,12aBC | 0,2±0,12abBC | 0,3±0,11abBC | 0,1±0,14abBC | 0,2±0,15abBC | 0,2±0,11abBC | ||
| 4 | 0,4±0,11abA | 0,4±0,12abA | 0,3±0,11bA | 0,3±0,14abA | 0,3±0,12abA | 0,4±0,11aA | 0,3±0,11abA | 0,3±0,11abA | 0,2±0,11abA | 0,4±0,10abA | 0,4±0,14abA | ||
| 5 | 0,2±0,10abAB | 0,1±0,12abAB | 0,1±0,11bAB | 0,2±0,13abAB | 0,2±0,10abAB | 0,4±0,09aAB | 0,4±0,14abAB | 0,4±0,10abAB | 0,3±0,12abAB | 0,1±0,16abAB | 0,5±0,10abAB | ||
| 6 | 0,1±0,10abBC | 0,1±0,11abBC | 0,1±0,12bBC | 0,1±0,10abBC | 0,2±0,11abBC | 0,3±0,14aBC | 0,2±0,11abBC | 0,2±0,12abBC | 0,2±0,11abBC | 0,3±0,12abBC | 0,3±0,09abBC | ||
| OPEFB | 2 | 0±0,11aB | 0±0,11aB | 0±0,10aB | 0±0,12aB | 0±0,11aB | 0±0,14aB | 0±0,11aB | 0±0,10aB | 0±0,12aB | 0±0,11aB | 0±0,15aB | |
| 3 | 0±0,11aB | 0±0,11aB | 0±0,14aB | 0±0,12aB | 0±0,10aB | 0±0,11aB | 0±0,11aB | 0±0,08aB | 0±0,10aB | 0±0,10aB | 0±0,11aB | ||
| 4 | 0±0,10aAB | 0±0,11aAB | 0,1±0,14aAB | 0±0,11aAB | 0±0,09aAB | 0±0,10aAB | 0±0,12aAB | 0,1±0,10aAB | 0±0,08aAB | 0,1±0,11aAB | 0±0,09aAB | ||
| 5 | 0±0,14aA | 0±0,11aA | 0,1±0,12aA | 0±0,10aA | 0±0,11aA | 0,1±0,10aA | 0,1±0,09aA | 0,1±0,10aA | 0,1±0,12aA | 0±0,11aA | 0,2±0,11aA | ||
| 6 | 0,1±0,11aA | 0±0,10aA | 0±0,11aA | 0,1±0,12aA | 0±0,10aA | 0±0,11aA | 0,1±0,11aA | 0,1±0,10aA | 0,1±0,12aA | 0,1±0,10aA | 0,2±0,11aA | ||
for xylitol production. The quantity of xylitol generated was contingent upon the xylose concentration in the medium; thus, ethanol was not detected in the medium, indicating the absence of fermentation (refer to Figure 3).
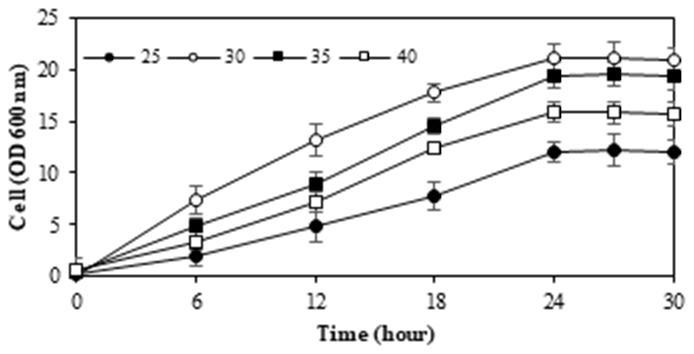
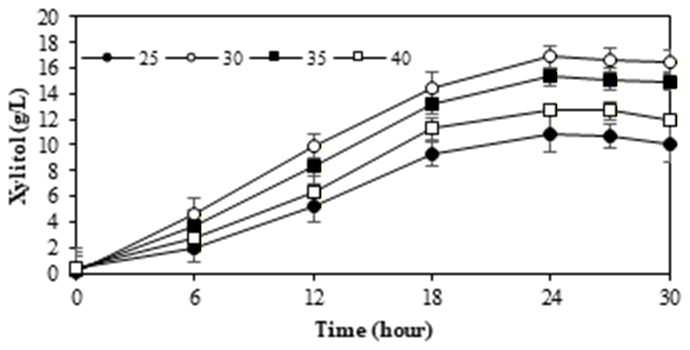
3.4. M. guilliermondii InaCCY65 in various pH and temperatures
To identify yeast species with superior xylitol production capabilities across a broad pH range (3.0-6.0) and temperature range (25-40°C), a cell growth assay was conducted using xylose, a commonly utilized medium. The performance of M. guilliermondii InaCCY65 across various pH levels and temperatures is depicted in Figure 4.
When cultivated in xylose, the optimal temperature for xylitol production by InaCCY65 was 30°C at pH 4.5, with diminished cell growth observed at both 25°C and 40°C.
The stationary phase of cell growth was obtained after 24 hours. Moreover, the ideal pH for cell growth and xylitol production by M. guilliermondii InaCCY65 was pH 4.5. Deviations from this pH value resulted in reduced efficacy. These findings align with previous results (refer to Figure 2 and Table 1), underscoring the ability of M. guilliermondii InaCCY65 to produce xylitol within the pH range of 3 to 5. Various reports have highlighted the influence of temperature conditions on xylitol production by different yeast strains. Optimal xylitol yields were achieved under specific conditions: Candida parapsilosis at pH 5.0; Candida guilliermondii at pH 4.5; Candida boidinii at pH 6.049); and Hansenula anomala50) at pH 4.551). Additionally, the pH range for xylose-to-xylitol bioconversion by Debaryomyces hansenii UFV-170 was



reported to be 4–851), while Pichia stipitis demonstrated optimal performance at pH 5.051). Kluyveromyces marxianus CCA510 exhibited peak activity at pH 6.052), and Candida boidinii was most effective at pH 6.053).
3.5. Characterization of selected xylitol-producing yeast in initial xylose concentration
The initial concentration of xylose as a substrate significantly influences xylitol fermentation. M. guilliermondii InaCCY65 was cultured in media containing xylose concentrations ranging from 10 to 80 g/L to investigate the impact of initial substrate concentration on xylitol production capacity. A summary of the experiments assessing the effect of xylose concentration on xylitol production by M. guilliermondii InaCCY65 is presented in Figure 5.
Figure 5 summarizes xylitol production by M. guilliermondii InaCCY65 across various initial concentrations of xylose. The Figure illustrates that xylitol production commences following the assimilation of xylose, with higher xylitol concentrations observed as the initial xylose concentration increases, peaking after 24 hours of incubation. Elevated xylose concentrations corresponded to increased xylitol levels. However, the yield coefficient (YP/S) after 24 hours of incubation at each initial xylose concentration was observed to decrease, with values of 0.71, 0.45, 0.42, 0.34, and 0.33, indicating a reduction in YP/S as xylose concentration rises. The highest YP/S of 0.71 was achieved at an initial xylose concentration of 20 g/L. These findings suggest an inhibitory effect of high xylose concentrations on the
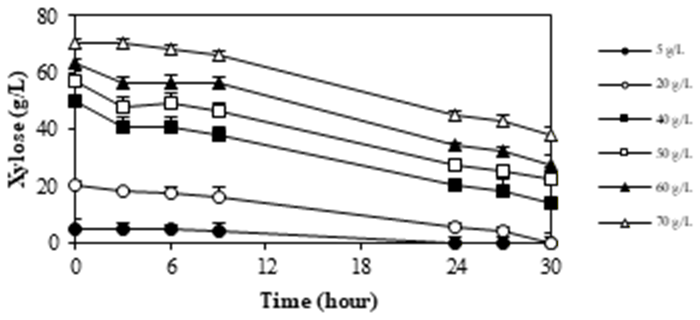
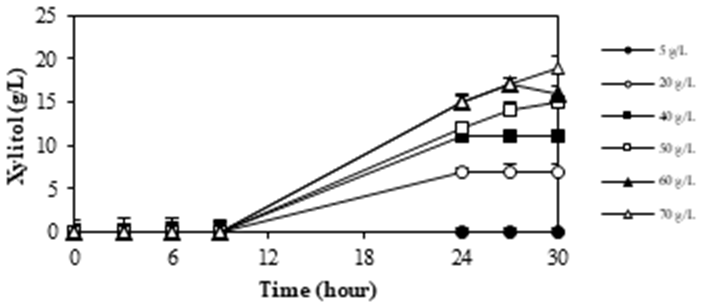
growth of InaCCY65, resulting in a slower attainment of xylitol conditions. Consequently, the results indicate that an initial xylose concentration of 20 g/L is the most efficient for xylitol production.
Deng et al. reported the influence of initial xylose on xylitol production by Wickerhamomyces anomalus WA found that increasing the xylose concentration from 20 g/L to 180 g/L resulted in a corresponding increase in xylitol production from 0.61 g/g to 0.91 g/g. These findings were attained by supplementing the fermentation with 20 g/L glycerol as a co-substrate23).
3.6. Growth profile of InaCCY65 in medium containing inhibitory chemical complex (ICC)
The main obstacle in utilizing agricultural by-products for xylitol production lies in the existence of ICCs54,55). To assess the impact of these inhibitors on xylitol production, M. guilliermondii InaCCY65 was cultured in media containing varying concentrations of ICCs. The concentration of ICCs in the growth medium was adjusted to achieve concentrations ranging from 5.0 to 70.0 mM furfural. The influence of ICCs on both cell growth and xylitol production is depicted in Figure 6.
The rate of cell growth and xylose consumption exhibits variability depending on the concentration of ICCs present in the medium. Xylitol production is closely linked to the growth rate, as it escalates promptly following the onset of xylose consumption. In the YPXi medium, the strain experiences minimal growth and fails to consume xylose. Essentially, when ICC concentrations are excessively high, xylose assimilation ceases, leading to a complete halt in cell growth.
Figure 6 depicts a delay in xylose consumption as ICC concentrations increase. The rate of xylose assimilation varies with ICC levels in the medium. Higher ICC concentrations prolong xylose assimilation during fermentation. In the 0.5YPX+0.5YPXi medium, a reduction in xylose assimilation is evident, with some remaining unused until the end of fermentation, after 30 hours of incubation. This slowdown in xylose consumption affects both cell growth and xylitol production. Similar effects are observed in the 0.25YPX+0.75YPXi and YPXi media. These results indicate that M. guillormondii InaCCY65 exhibits resilience to inhibitory compounds, tolerating ICC concentrations up to 75% of the total. Despite significant inhibition of cell growth in the YPXi medium, InaCCY65 cells fail to assimilate xylose, resulting in reduced xylitol production, approximately around 2 g/L. Furfural toxicity has been associated with modifications in cell wall and plasma membrane composition, DNA damage, oxidative stress, suppression of RNA synthesis, and disruption of protein synthesis56).
Additionally, it induces the aggregation of tubular mitochondria and inhibits enzymes involved in central carbon metabolism57). The compound of 5-HMF is hazardous because it inhibits glycolysis and disrupts RNA and protein synthesis58). Furthermore, inhibitor compounds have been documented to impede xylose reductase activity in the xylose metabolism of Thermomyces lanuginosus59) and D. nepalensis60). The cell growth and xylitol production of these strains inhibited in the presence of furfural, 5-HMF, acetic acid, vanillin, gallic acid, and phenol.
Several approaches can be employed to mitigate the toxicity of lignocellulosic hydrolysates to yeast, including hydrolysate detoxification, cell encapsulation, cultivation with high cell density59), adaptive strain development33), and genetic engineering61). However, these strategies inevitably entail increased energy consumption and costs. The inherent resistance of M. guilliermondii InaCCY65 to inhibitors presents a promising avenue for its utilization as a cell factory in xylitol fermentation from lignocellulosic hydrolysates.
3.7. The profiles of detoxification of inhibitory chemical complex (ICC)
Superior isolates may employ several physiological mechanisms to reduce the effects of ICC compounds on cell growth and their ability to produce xylitol. Therefore, the ability to grow and produce xylitol of strain InaCCY65 was tested using different concentrations of ICCs, including 5-HMF, furfural, formic acid, acetic acid and levulinic acid. The detoxification of the tested compounds by the InaCCY65 strain is shown in Figure 7.
Based on the cell growth profile and inhibitor detoxification, the cells exhibited survival through successful detoxification of inhibitors, namely 5-HMF and furfural (as depicted in Figures 7A and 7B).




The concentrations of furfural and 5-HMF declined during the fermentation process in accordance with their initial concentrations. In the YPXi medium, cell death occurred due to an inability to withstand the inhibitors, as indicated by the inhibitor profiles showing unsuccessful detoxification. In this medium, furfural remained at its original concentration of 40%, while 5-HMF was less

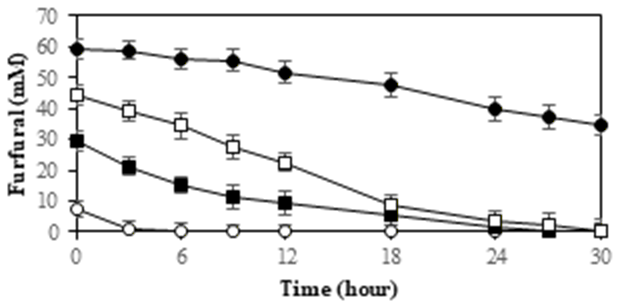

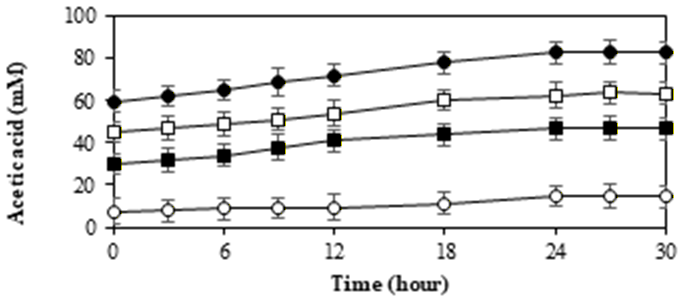

detoxified, with 80% of its original concentration remaining due; to We observed that InaCCY65 cells managed to detoxify 5-HMF and furfural as the concentrations of inhibitors decreased during fermentation, particularly in the 0.75YP+0.25YPi, 0.5YP+0.5YPi, and 0.25YP+0.75YPi media. It is believed that 5-HMF and furfural directly inhibit the activity of critical enzymes involved in xylose metabolism, such as xylose reductase62). An intriguing aspect that warrants further investigation is the bioconversion of furfural and 5-HMF compounds into other compounds, which can fulfilled the information of their metabolism.
4. Conclusion
In conclusion, Meyerozyma guilliermondii InaCCY65 excels as a top candidate for converting lignocellulosic biomass into xylitol due to its adaptability and productivity. Thriving in diverse hydrolysates and acidic conditions, the strains of InaCCY65 consistently produce xylitol and utilize both glucose and xylose efficiently. The optimal conditions of temperature and pH were 30°C and pH 4.5 to further enhance its productivity. The InaCCY65's resilience against inhibitors and acidic environments were ideal for industrial applications, promising innovations in sustainable xylitol production and bioeconomy advancement.
Acknowledgements
This work was supported by the National Research and Innovation Agency (BRIN) [RP3_Tema2_036, 2023-2024). I am deeply thankful to Prof. Dr. I Made Sudiana, M.Sc for his insightful discussions, suggestion and moral support. I dedicated this paper for the mentorship of the late Prof. Dr. Ir. Euis Hermiati, M.Sc and I would like to express my most profound respect for her kindly support until this paper published.
References
- N. Dragomir, D.M. Grigore, and E.N. Pogurschi, “Beyond sugar: a holistic review of sweeteners and their role in modern nutrition,” Foods, 14 (18) 3182 (2025). doi:10.3390/foods14183182.
- F. Cocco, M.G. Cagetti, O. Majdub, and G. Campus, “Concentration in saliva and antibacterial effect of xylitol chewing gum: in vivo and in vitro study,” Applied Sciences, 10 (8) 2900 (2020). doi:10.3390/app10082900.
- N.Z. Msomi, O.L. Erukainure, and M.S. Islam, “Suitability of sugar alcohols as antidiabetic supplements: a review,” Journal of Food and Drug Analysis, 29 (1), 1–14 (2021). doi:10.38212/2224-6614.3107.
- N.M. Clauser, C.G. Fit, R.E. Cardozo, J.A. Rivaldi, F.E. Felissia, M.C. Area, and M.E. Vallejos, “Technical, economic and environmental assessment of xylitol production in a biorefinery platform toward a circular economy,” Sustainability, 16 (23) 10770 (2024). doi:10.3390/su162310770.
- L. Wu, X. Ma, X. Liu, H. Du, and F. Wang, “A phyllosilicate-derived Cu–Ni alloy catalyst for efficient hydrogenation of high-concentration xylose aqueous solution to xylitol,” Industrial and Engineering Chemistry Research, 64 (22), 10740–10748 (2025). doi:10.1021/acs.iecr.5c00713.
- D.J. Aigner, L. Hinterholzer, L. Almhofer, R.H. Bischof, and T. Wrodnigg, “Conversion of xylose into D-xylitol using catalytic transfer hydrogenation with formic acid as H-donor,” BioResources, 18 (4), 8631–8652 (2023). doi:10.15376/biores.18.4.8631-8652.
- H.B. Du, X.P. Ma, M. Jiang, S. Zhao, Q. Fang, X. Liu, and Z.C. Zhang, “Xylitol production from xylose by catalytic hydrogenation over an efficient Cu–Ni/SiO2 bimetallic catalyst,” ACS Sustainable Chemistry & Engineering, 11 (6), 2115–2126 (2023). doi:10.1021/acssuschemeng.2c04964.
- W. Yan, H. Gao, X. Qian, Y. Jiang, J. Zhou, W. Dong, F. Xin, W. Zhang, and M. Jiang, “Biotechnological applications of the non-conventional yeast Meyerozyma guilliermondii,” Biotechnology Advances, 46 107674 (2021). doi:10.1016/j.biotechadv.2020.107674.
- H.Y. Leong, C.K. Chang, K.S. Khoo, K.W. Chew, S.R. Chia, J.W. Lim, J.S. Chang, and P.L. Show, “Waste biorefinery towards a sustainable circular bioeconomy: a solution to global issues,” Biotechnology for Biofuels, 14 (1), 1–15 (2021). doi:10.1186/s13068-021-01939-5.
- A. Pandey, and Y.C. Sharma, “Advancements in biomass valorization in integrated biorefinery systems,” Biofuels, Bioproducts and Biorefining, 18 (6) (2024). doi:10.1002/bbb.2670.
- S. Steven, N.T.U. Culsum, I.C. Sophiana, I. Febijanto, E. Syamsudin, N. Ghazali, N. Nadirah, E.S.A. Soekotjo, and I.M. Hidayatullah, “Potential of corn cob sustainable valorization to fuel-grade bioethanol: a simulation study using SuperPro Designer®,” EVERGREEN – Joint Journal of Novel Carbon Resource Sciences & Green Asia Strategy, 10 (4), 2287–2298 (2023). doi:10.5109/7160904.
- A. Singh, K. Kanchan, Sadhvi, M. Saxena, and N. Sharma, “Production process of sugarcane bagasse for the generation of bio-products and biofuel: an overview,” Archives of Current Research International, 24 (12), 231–240 (2024). doi:10.9734/acri/2024/v24i121014.
- S. Singh, L. Nagdeve, H. Kumar, and K. Dhakar, “Rice straw based natural fiber reinforced polymer for sustainable bio-composites: a systematic review,” EVERGREEN – Joint Journal of Novel Carbon Resource Sciences & Green Asia Strategy, 10 (2),1041–1052 (2023). doi:10.5109/6793661.
- A. Liu, Y. Zhao, S. Wang, X. Jiang, and Q. Huang, “Utilization and preparation strategies of biomass containing lignin and cellulose: a review,” Materials Today Chemistry, 48 102926 (2025). doi:10.1016/j.mtchem.2025.102926.
- X. Peng, S. Nie, X. Li, X. Huang, and Q. Li, “Characteristics of the water- and alkali-soluble hemicelluloses fractionated by sequential acidification and graded-ethanol from sweet maize stems,” Molecules, 24 (1) 212 (2019). doi:10.3390/molecules24010212.
- M. Tišma, A. Bušić-Kojić, and M. Planinić, “Bio-based products from lignocellulosic waste biomass: a state of the art,” Chemical and Biochemical Engineering Quarterly, 35 (2), 139–156 (2021). doi:10.15255/CABEQ.2021.1931.
- M.A. Lorenzo-Santiago, J. Rodríguez-Campos, R. Rendón-Villalobos, E. García-Hernández, A.A. Vallejo-Cardona, and S.M. Contreras-Ramos, “Thermal treatment to obtain 5-hydroxymethyl furfural (5-HMF), furfural and phenolic compounds from vinasse waste from agave,” Molecules, 28 (3) 1063 (2023). doi:10.3390/molecules28031063.
- T.M. Louie, K.C. Louie, S. DenHartog, S. Gopishetty, M. Subramanian, M.A. Arnold, and S. Das, “Production of bio-xylitol from D-xylose by an engineered Pichia pastoris expressing a recombinant xylose reductase without auxiliary substrate as electron donor,” Microbial Cell Factories, 20 (1) 50 (2021). doi:10.1186/s12934-021-01534-1.
- V. Tadioto, L.M. Milani, É.T. Barrilli, C.W. Baptista, L. Bohn, A. Dresch, R. Harakava, O. Fogolari, G.M. Mibielli, J.P. Bender, H. Treichel, B.U. Stambuk, C. Müller, and S.A. Alves Jr, “Analysis of glucose and xylose metabolism in new indigenous Meyerozyma caribbica strains isolated from corn residues,” World Journal of Microbiology and Biotechnology, 38 (2) Article 22 (2022). doi:10.1007/s11274-021-03221-0.
- J. Montaño López, L. Duran, and J.L. Avalos, “Physiological limitations and opportunities in microbial metabolic engineering,” Nature Reviews Microbiology, 20 (1), 35–48 (2022). doi:10.1038/s41579-021-00600-0.
- G.C. Bevilaqua, F. Maugeri, and M.B.S. Forte, “Simultaneous production of xylitol and arabitol by Candida tropicalis fermentation improving agro-industrial wastes valorization,” Food and Bioproducts Processing, 140, 29–45 (2023). doi:10.1016/j.fbp.2023.04.006.
- S. Pant, R. Ritika, A. Prakash, and A. Kuila, “Integrated production of ethanol and xylitol from Brassica juncea using Candida sojae JCM 1644,” Bioresource Technology, 351 126903 (2022). doi:10.1016/j.biortech.2022.126903.
- Y. Deng, X. Luo, H. Wang, S. Li, J. Liang, and Z. Pang, “Xylitol fermentation characteristics with a newly isolated yeast Wickerhamomyces anomalus WA,” Fungal Biology, 128 (2), 1657–1663 (2024). doi:10.1016/j.funbio.2024.01.004.
- N. Hidayah, R.S. Adiandri, and E. Rahayu, “Microbial xylitol production from corn cob using Candida guilliermondii,” IOP Conference Series: Earth and Environmental Science, 1024 (1) 012077 (2022). doi:10.1088/1755-1315/1024/1/012077.
- S. Singh, and S.N. Gummadi, “Two-stage seeding strategy and its multi-response optimization for enhanced xylitol production by Debaryomyces nepalensis NCYC 3413,” Bioresource Technology, 413 131469 (2024). doi:10.1016/j.biortech.2024.131469.
- S.F.A. Manaf, A.A.I. Luthfi, N.Z. Nasoha, N.S. Engliman, N.S. Jamali, and T.P. Tan, “Phenotype adaptation of Kluyveromyces marxianus for enhanced conversion of biomass into xylitol,” Process Biochemistry, 139, 104–115 (2024). doi:10.1016/j.procbio.2024.01.019.
- F.A. da Silveira, T.A.R. Fernandes, C.R.S. Bragança, T.R. Balbino, R.H.S. Diniz, F.M.L. Passos, and W.B. da Silveira, “Isolation of xylose-assimilating yeasts and optimization of xylitol production by a new Meyerozyma guilliermondii strain,” International Microbiology, 23 (2), 325–334 (2020). doi:10.1007/s10123-019-00105-0.
- A. Ochoa-Chacón, A.C. Ramos-Valdivia, H.M. Poggi-Varaldo, L. Villa-Tanaca, A. Martinez, and T. Ponce-Noyola, “Fermentation performance of a Mexican native Clavispora lusitaniae strain for xylitol and ethanol production from xylose, glucose and cellobiose,” Enzyme and Microbial Technology, 160 110094 (2022). doi:10.1016/j.enzmictec.2022.110094.
- H.F. Son, S. Lee, and K. Kim, “Structural insight into D-xylose utilization by xylose reductase from Scheffersomyces stipites,” Scientific Reports, 8, 1–11 (2018). doi:10.1038/s41598-018-35703-x.
- E. Hermiati, R.P.B. Laksana, W. Fatriasari, L.N. Kholida, A. Thontowi, Yopi, D.R. Arnieyanto, V. Champreda, and T. Watanabe, “Microwave-assisted acid pretreatment for enhancing enzymatic saccharification of sugarcane trash,” Biomass Conversion and Biorefinery, 13, 3037–3047 (2023). doi:10.1007/s13399-020-00971-z.
- D. Umai, R. Kayalvizhi, V. Kumar, and S. Jacob, “Xylitol: bioproduction and applications – a review,” Frontiers in Sustainability, 3 826190 (2022). doi:10.3389/frsus.2022.826190.
- V.C. Ujor, and C.C. Okonkwo, “Microbial detoxification of lignocellulosic biomass hydrolysates: biochemical and molecular aspects, challenges, exploits and future perspectives,” Frontiers in Bioengineering and Biotechnology, 10 1061667 (2022). doi:10.3389/fbioe.2022.1061667.
- S. Kaur, P. Guleria, and S.K. Yadav, “Evaluation of fermentative xylitol production potential of adapted strains of Meyerozyma caribbica and Candida tropicalis from rice straw hemicellulosic hydrolysate,” Fermentation, 9 (2), 181 (2023). doi:10.3390/fermentation9020181.
- L.M. Eraso, O. Cuaspud, and M.E. Arias Zabala, “Optimization of xylitol production through Candida tropicalis in xylose hydrolysate from rice husk,” Biomass Conversion and Biorefinery, 15, 26085–26095 (2025). doi:10.1007/s13399-024-05372-0.
- P. Surain, and N.K. Aggarwal, “Candida, a human pathogen and major types of candidiasis,” International Journal of Pharmaceutical Sciences and Research, 11 (1), 41–67 (2020). doi:10.13040/IJPSR.0975-8232.11(1).41-67.
- N. Liu, C. Dou, X. Yang, B. Bai, S. Zhu, J. Tian, Z. Wang, L. Xu, and B. Shen, “Effects of pretreatment procedure, compositional feature and reaction condition on the devolatilization characteristics of biomass during pyrolysis process: a review,” Journal of the Energy Institute, 118 101943 (2025). doi:10.1016/j.joei.2024.101943.
- C.K. Yamakawa, L. Kastell, M.R. Mahler, J.L. Martinez, and S.I. Mussatto, “Exploiting new biorefinery models using non-conventional yeasts and their implications for sustainability,” Bioresource Technology, 309 123374 (2020). doi:10.1016/j.biortech.2020.123374.
- P. Kahar, E.I. Riyanti, H. Otsuka, H. Matsumoto, C. Kihira, C. Ogino, and A. Kondo, “Challenges of non-flocculating Saccharomyces cerevisiae haploid strain against inhibitory chemical complex for ethanol production,” Bioresource Technology, 245, 1436–1446 (2017). doi:10.1016/j.biortech.2017.06.009.
- C.E. O’Brien, J. Oliveira-Pacheco, E.O. Cinnéide, M.A.B. Haase, C.T. Hittinger, T.R. Rogers, O. Zaragoza, U. Bond, and G. Butler, “Population genomics of the pathogenic yeast Candida tropicalis identifies hybrid isolates in environmental samples,” PLoS Pathogens, 17 (3) e1009138 (2021). doi:10.1371/journal.ppat.1009138.
- P. Bonthong, B. Bunterngsook, W. Mhuantong, K. Aiewviriyasakul, W. Sritusnee, V. Champreda, and H. Lekakarn, “Genomic and functional analysis of a novel yeast Cyberlindnera fabianii TBRC 4498 for high-yield xylitol production,” Journal of Fungi, 11 (6), 453 (2025). doi:10.3390/jof11060453.
- F. Palladino, R.C.L.B. Rodrigues, R.M. Cadete, K.O. Barros, and C.A. Rosa, “Novel potential yeast strains for the biotechnological production of xylitol from sugarcane bagasse,” Biofuels, Bioproducts and Biorefining, 15 (3), 690–702 (2021). doi:10.1002/bbb.2196.
- C.V.G.C. Carneiro, F.C.P. Silva, and J.R.M. Almeida, “Xylitol production: identification and comparison of new producing yeasts,” Microorganisms, 7 (11), 484 (2019). doi:10.3390/microorganisms7110484.
- R.D. Umai, S. Jacob, and V. Kumar, “Deep eutectic solvent pretreatment of water hyacinth for improved holocellulosic saccharification and fermentative co-production of xylitol and lipids using Rhodosporidium toruloides NCIM 3547,” Fermentation, 8 (11), 591 (2022). doi:10.3390/fermentation8110591.
- S. Singh, S.K. Arya, and M. Krishania, “Bioprocess optimization for enhanced xylitol synthesis by new isolate Meyerozyma caribbica CP02 using rice straw,” Biotechnology for Biofuels and Bioproducts, 17, 31 (2024). doi:10.1186/s13068-024-02475-8.
- B.R.A. Alencar, R.A.A. Freitas, V.E.P.G. Silva, R.K. Silva, C. Elsztein, S.P. Silva, E.D. Dutra, M.A. Morais Junior, and R.B. Souza, “Meyerozyma caribbica isolated from vinasse-irrigated sugarcane plantation soil: a promising yeast for ethanol and xylitol production in biorefineries,” Journal of Fungi, 9 (8), 789 (2023). doi:10.3390/jof9080789.
- D. Dasgupta, A. Sidana, B. Sarkar, S. More, D. Ghosh, T. Bhaskar, and A. Ray, “Process development for crystalline xylitol production from corncob biomass by Pichia caribbica,” Food and Bioproducts Processing, 133, 45–56 (2022). doi:10.1016/j.fbp.2022.02.006.
- V. Narisetty, E. Castro, S. Durgapal, F. Coulon, S. Jacob, D. Kumar, M.K. Awasthi, K.K. Pant, B. Parameswaran, and V. Kumar, “High level xylitol production by Pichia fermentans using non-detoxified xylose-rich sugarcane bagasse and olive pits hydrolysates,” Bioresource Technology, 342 126005 (2021). doi:10.1016/j.biortech.2021.126005.
- E. Mardawati, D.N. Daulay, D.W. Wira, and E. Sukarminah, “The effect of initial cell and pH on xylitol fermentation from oil palm empty fruit bunch,” Industria: Journal of Agro-Industrial Management Technology, 7 (1), 23–30 (2018). doi:10.21776/ub.industria.2018.007.01.3.
- S. Bedő, A. Fehér, P. Khunnonkwao, K. Jantama, and C. Fehér, “Optimized bioconversion of xylose derived from pre-treated crop residues into xylitol by using Candida boidinii,” Agronomy, 11 (1), 79 (2021). doi:10.3390/agronomy11010079.
- A.M. Erian, and M. Sauer, “Utilizing yeasts for the conversion of renewable feedstocks to sugar alcohols – a review,” Bioresource Technology, 346 126296 (2022). doi:10.1016/j.biortech.2021.126296.
- A. Bhattacharya, A. Ganguly, A.K. Sadhukhan, and P.K. Chatterjee, “Investigations on the effect of driving parameters for xylitol production from water hyacinth biomass,” Indian Journal of Biotechnology, 17 (2), 272–283 (2018).
- T.L. Albuquerque, S.D.L. Gomes, J.E. Marques, I.J. Silva, and M.V.P. Rocha, “Xylitol production from cashew apple bagasse by Kluyveromyces marxianus CCA510,” Catalysis Today, 255, 33–40 (2015). doi:10.1016/j.cattod.2014.10.054.
- S.F.A. Manaf, A.A.I. Luthfi, J.M. Jahim, S. Harun, J.P. Tan, and S.H.M. Shah, “Sequential detoxification of oil palm fronds hydrolysate with coconut shell activated charcoal and pH controlled in bioreactor for xylitol production,” Chemical Engineering Research and Design, 179, 90–106 (2022). doi:10.1016/j.cherd.2022.01.008.
- R. Sharma, D. Vaidya, M. Kaushal, A. Gupta, Shavnam, and C. Thakur, “Valorization of corn waste for hemicellulose extraction and xylitol production: comparative study of different extraction methods,” Biofuels, Bioproducts and Biorefining, 19, 1746–1764 (2025). doi:10.1002/bbb.2782.
- I.A. Bianchini, F.M. Jofre, S.S. Queiroz, T.M. Lacerda, and M.G.A. Felipe, “Relation of xylitol formation and lignocellulose degradation in yeast,” Applied Microbiology and Biotechnology, 107 (10), 3143–3151 (2023). doi:10.1007/s00253-023-12495-3.
- L. Wang, A. Qi, J. Liu, Y. Shen, and J. Wang, “Comparative metabolic analysis of the adaptive Candida tropicalis to furfural stress response,” Chemical Engineering Science, 267 118348 (2023). doi:10.1016/j.ces.2022.118348.
- M. Knesebeck, M. Rüllke, V. Schönrock, J.P. Benz, and D. Weuster-Botz, “D-xylitol production from sugar beet press pulp hydrolysate with engineered Aspergillus niger,” Microorganisms, 12 (12) 2489 (2024). doi:10.3390/microorganisms12122489.
- E. Gencturk, and K.O. Ulgen, “Understanding HMF inhibition on yeast growth coupled with ethanol production for the improvement of bio-based industrial processes,” Process Biochemistry, 121, 425–438 (2022). doi:10.1016/j.procbio.2022.07.015.
- M. Zhang, A.K. Puri, Z. Wang, S. Singh, and K. Permaul, “A unique xylose reductase from Thermomyces lanuginosus: effect of lignocellulosic substrates and inhibitors and applicability in lignocellulosic bioconversion,” Bioresource Technology, 281, 374–381 (2019). doi:10.1016/j.biortech.2019.02.102.
- A. Paidimuddala, A. Rathod, and S.N. Gummadi, “Inhibition of Debaryomyces nepalensis xylose reductase by lignocellulose-derived by-products,” Biochemical Engineering Journal, 121, 73–82 (2017). doi:10.1016/j.bej.2017.01.019.
- A.K. Singh, F. Deeba, M. Kumar, S. Kumari, S.A. Wani, T. Paul, and N.A. Gaur, “Development of engineered Candida tropicalis strain for efficient corncob-based xylitol-ethanol biorefinery,” Microbial Cell Factories, 22 (1), 201 (2023). doi:10.1186/s12934-023-02190-3.
- Y. Lugani, M. Puri, and B.S. Sooch, “Recent insights, applications and prospects of xylose reductase: a futuristic enzyme for xylitol production,” European Food Research and Technology, 247 (4), 921–946 (2021). doi:10.1007/s00217-020-03674-x.